| 1. 染色体の示す柔軟な「再編成」能力への実験的介入 |
|
|
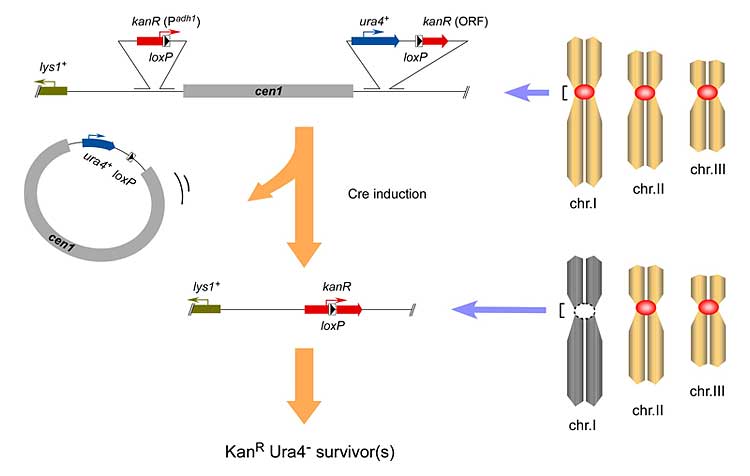
 各々の細胞に含まれる長大なDNA分子は遺伝情報の担い手であり、染色体はそのようなDNAを安定に維持・継承し、その機能発揮を補完する構造体として発達してきました。数々の細胞分裂を経て起こる個体発生の過程でも、個体間の生殖を介して起こる子孫形成の過程でも、染色体はただひたすらにDNAの恒常性を精巧に担い続けています。しかしながら種分化や生物進化の時間軸で染色体を捉えると、そのDNA構造はむしろ変化を繰り返しているように映ります。すなわち、染色体転座などによる折々のDNA恒常性破綻に際して、染色体は甘んじて死を受け入れるのではなく、自らを再編成することで破綻に順応し、再び恒常性ある構造体に回復していることが示唆されるのです。その成立こそが種分岐の契機である可能性があります。私たちは研究モデルである分裂酵母に染色体変化を強制する実験系を確立し、柔軟な染色体再編成の分子基盤と制御機構について直接的に解析を行います。 各々の細胞に含まれる長大なDNA分子は遺伝情報の担い手であり、染色体はそのようなDNAを安定に維持・継承し、その機能発揮を補完する構造体として発達してきました。数々の細胞分裂を経て起こる個体発生の過程でも、個体間の生殖を介して起こる子孫形成の過程でも、染色体はただひたすらにDNAの恒常性を精巧に担い続けています。しかしながら種分化や生物進化の時間軸で染色体を捉えると、そのDNA構造はむしろ変化を繰り返しているように映ります。すなわち、染色体転座などによる折々のDNA恒常性破綻に際して、染色体は甘んじて死を受け入れるのではなく、自らを再編成することで破綻に順応し、再び恒常性ある構造体に回復していることが示唆されるのです。その成立こそが種分岐の契機である可能性があります。私たちは研究モデルである分裂酵母に染色体変化を強制する実験系を確立し、柔軟な染色体再編成の分子基盤と制御機構について直接的に解析を行います。
| 2. 染色体に宿った「外来」RNA要素に対する対処の理解 |
|
|
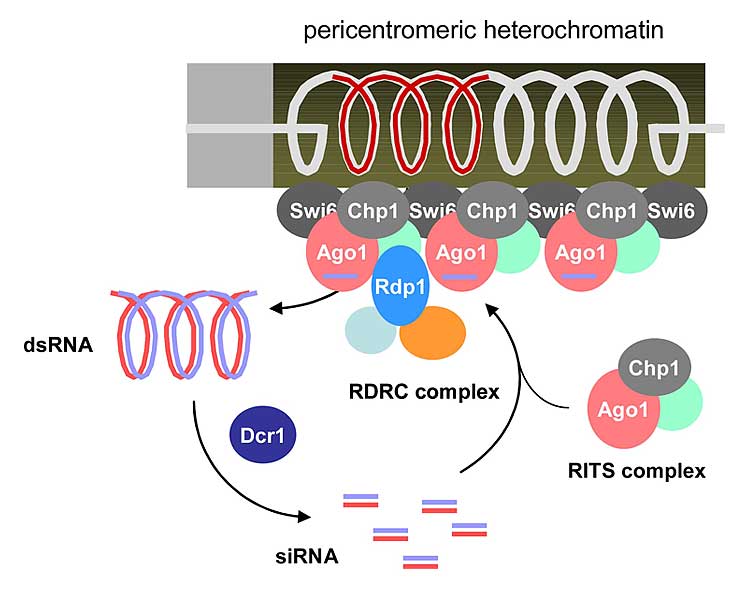
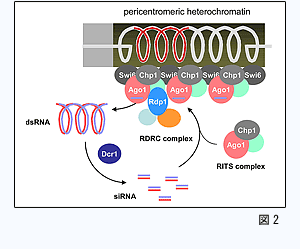 染色体DNAの恒常性を脅かす要素の一つに、細胞外来のRNA因子の侵入があります。そのゲノムDNAへの反復した挿入は、挿入部位によって遺伝子機能を阻害する可能性があるだけでなく、ゲノムDNA構造そのものを不安定化させる危険性があります。しかし、一度染色体に挿入された外来RNA因子を内在性のRNAと区別することは容易ではありません。分裂酵母ではヘテロクロマチン構造を形成する反復DNA領域から非コードRNAが生成され、それはRNAi機構の標的となることが知られています。そのような制御が働くのは、反復DNA由来の非コードRNAがかつて外来因子として排除されていた名残である可能性が考えられます。私たちは、RNAi標的化機構の解析を通じて真の内在性RNAと見かけ上内在の外来性RNA要素を区別する仕組みの一端を求めるとともに、ヘテロクロマチン形成への関連の追究を通じてその制御の道理と意義を探ります。 染色体DNAの恒常性を脅かす要素の一つに、細胞外来のRNA因子の侵入があります。そのゲノムDNAへの反復した挿入は、挿入部位によって遺伝子機能を阻害する可能性があるだけでなく、ゲノムDNA構造そのものを不安定化させる危険性があります。しかし、一度染色体に挿入された外来RNA因子を内在性のRNAと区別することは容易ではありません。分裂酵母ではヘテロクロマチン構造を形成する反復DNA領域から非コードRNAが生成され、それはRNAi機構の標的となることが知られています。そのような制御が働くのは、反復DNA由来の非コードRNAがかつて外来因子として排除されていた名残である可能性が考えられます。私たちは、RNAi標的化機構の解析を通じて真の内在性RNAと見かけ上内在の外来性RNA要素を区別する仕組みの一端を求めるとともに、ヘテロクロマチン形成への関連の追究を通じてその制御の道理と意義を探ります。
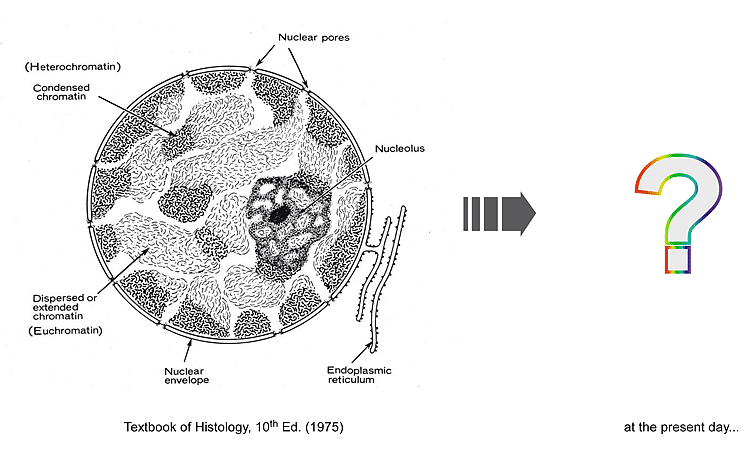
 真核生物細胞では、DNAを機能的に収納する構造体として染色体に加えて細胞核が挙げられます。染色体は一本のDNA分子から形成されますが、ゲノムとは複数のDNA分子で成り立つものであり、細胞核は複数の染色体に分割されたゲノムを総括して包含する役目を細胞内器官として担っています。染色体の大規模な「再編成」能力やトランスに働く「外来」RNA要素の制御には、細胞核のような高次レベルでの秩序が高く関与することは間違いありません。しかしながら、その実体はまだ不明瞭なままです。またそのような秩序は、ゲノムDNAからの遺伝子発現を介した生命プログラムの発動にも大きく関わるものと考えられます。私たちは適切な遺伝学アッセイに基づく解析ツールを分裂酵母に設定し、ヘテロクロマチン構造やRNAi機構がいかなる仕組みで細胞核の特定の空間領域に画定されるのか、という課題にも取り組みます。そのような「構造的秩序」の解明は、染色体「再編成」や「外来」因子の制御への理解にも役立つものと考えます。 真核生物細胞では、DNAを機能的に収納する構造体として染色体に加えて細胞核が挙げられます。染色体は一本のDNA分子から形成されますが、ゲノムとは複数のDNA分子で成り立つものであり、細胞核は複数の染色体に分割されたゲノムを総括して包含する役目を細胞内器官として担っています。染色体の大規模な「再編成」能力やトランスに働く「外来」RNA要素の制御には、細胞核のような高次レベルでの秩序が高く関与することは間違いありません。しかしながら、その実体はまだ不明瞭なままです。またそのような秩序は、ゲノムDNAからの遺伝子発現を介した生命プログラムの発動にも大きく関わるものと考えられます。私たちは適切な遺伝学アッセイに基づく解析ツールを分裂酵母に設定し、ヘテロクロマチン構造やRNAi機構がいかなる仕組みで細胞核の特定の空間領域に画定されるのか、という課題にも取り組みます。そのような「構造的秩序」の解明は、染色体「再編成」や「外来」因子の制御への理解にも役立つものと考えます。
|